1
Anti-H3 | Histone H3 (rabbit antibody) (nuclear marker)
AS10 710 | Clonality: Polyclonal | Host: Rabbit | Reactivity: A. thaliana, B. cinerea, C. reinhardtii, G. hirsutum, H. vulgare, S. lycopersicum, V. faba, P. tricornutum, P. patens, S. europaea, Z. mays | cellular [compartment marker] of nucleoplasm
- Product Info
-
Immunogen: KLH-conjugated synthetic peptide derived from known H3 sequences, inluding Arabidopsis thaliana H3.3 P59169 (At4g40030, At4g40040, At5g10980), H3.2 P59226 (At1g09200, At3g27360, At5g10390, At5g10400, At5g65360), H3-like 2 Q9FXI7 (At1g19890)
Host: Rabbit Clonality: Polyclonal Purity: Serum Format: Lyophilized Quantity: 50 µl Reconstitution: For reconstitution add 50 µl of sterile water Storage: Store lyophilized/reconstituted at -20°C; once reconstituted make aliquots to avoid repeated freeze-thaw cycles. Please remember to spin the tubes briefly prior to opening them to avoid any losses that might occur from material adhering to the cap or sides of the tube. Tested applications: ChIp-qPCR (ChIp-qPCR), Immunocytochemistry (ICC), Immunofluorescence (IF), Western blot (WB) Recommended dilution: 1 : 100-1 : 500 (ICC), 2 µl of antibody/500 µl solution (ChIp-qPCR), 1: 500 (IF), 1 : 5000 (WB) Expected | apparent MW: 15 | 17 kDa - Reactivity
-
Confirmed reactivity: Arabidopsis thaliana, Botrytis cinerea, Brassica oleracea, Camellia sinensis, Capsicum annuum, Cicer arietinum L., Chlamydomonas acidophila, Chlamydomonas reinhardtii, Cucumis sativus L cv Suyo, Cucurbita pepo L. var. cylindrica, Hordeum vulgare, human, Nicotiana benthamiana, Oryza sativa, Phalaenopsis equestris, Phaeodactylum tricornutum, Physcomitrium patens, Salicornia europaea, Stevia rebaudiana, Solanum lycopersicum, Solanum sogarandinum, Solanum tuberosum, Vicia faba, Zea mays Predicted reactivity: Brachypodium distachyon, Brassica napus, Cyanidioschyzon merolae, Emiliania huxleyi, Fagopyrum tataricum Gaertn., Galdieria sulphuraria, Glycine max, Lolium multiflorum, Nannochloropsis gaditana, Nicotiana tabacum, Malus domestica, Medicago sativa, Oryza sativa, Ostreococcus sp. , Pinus pinaster, Pisum sativum, Populus sp., Porphyridium purpureum, Saccharum sp., Sorghum bicolor, Triticum aestivum, Vitis vinifera, Volvox sp.
Species of your interest not listed? Contact usNot reactive in: Cyanophora paradoxa - Application Examples
-
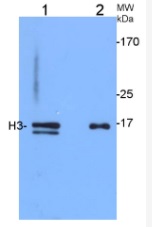
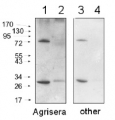
1.2 μg of Arabidopsis thaliana chromatin-enriched fraction (1) and 3.75 µg of total protein from 4-weeks-old Arabidopsis thaliana leaves (2), and were separated on 12% SDS-PAGE and blotted 50 mins to Immobilon-P (Millipore, semi-dry) PVDF membrane. Blots were blocked immediately following transfer in MTBS-T (5% milk) for 30 mins at room temperature with agitation. Blots were incubated in the primary antibody at a dilution of 1:5000 for 1h at room temperature with agitation. The antibody solution was decanted and the blot was rinsed briefly twice, then washed 3 times for 3 min in TBS-T at room temperature with agitation. Blots were incubated in secondary antibody (anti- IgG horse radish peroxidase conjugated, from Agrisera, AS09 602) diluted to 1:20 000 for 30 mins at room temperature with agitation. The blots were washed as above and developed for 5 min with ECL detection reagent according to the manufacturers instructions. Exposure time was 30 seconds. Double band in chromatine-enriched fraction (1) has been outcompeted in peptide neutralization assay by peptide used to elicit H3 antibodies. Chromatin izolation was carried out as described (Zilberman et al. 2008) with minor modifications.
Courtesy of Weronika Sura and Dr. Piotr A. Ziolkowski, Department of Biotechnology, Adam Mickiewicz University, Poznan, Poland

30 μg of 5 µl of Chlamydomonas reinhardtii protein saturated in 8M urea were separated on 15% SDS-PAGE and blotted for 1hour to 0.2 µm nitrocellulose at 100V using wet transfer system. Blots were blocked with 0.5% cold fish gelatin for 1hr at room temp with agitation. Blot was incubated in the primary antibody (anti-H3) at a dilution of 1:2500 for an hour at RT with agitation. The blots were washed with 3X 15min TBS-TT at RT with agitation. Blots as incubated in the secondary antibody (DayLight 800) 1:5000 dilution for 30min at RT with agitation and washed 1X with TBSTT for 15min, 1X with TBST for 15min before scanning with the ODyssey IRD scanner.
Courtesy of Dr. Betty Chung, University of Cambridge, United Kingdom 5 µl of 15μg/µl Solanum lycopersicum protein saturated in 8M urea were separated on 15% SDS-PAGE and blotted for 1hour to 0.2 µm nitrocellulose at 100V using wet transfer system. Blots were blocked with 0.5% cold fish gelatin for 1hr at room temp with agitation. Blot was incubated in the primary antibody (anti-H3) at a dilution of 1:2500 for an hour at RT with agitation. The blots were washed with 3X 15min TBS-TT at RT with agitation. Blots as incubated in the secondary antibody, fluorescent antibody (AS12 2460, Agrisera) 1:5000 dilution for 30min at RT with agitation and washed 1X with TBSTT for 15min, 1X with TBST for 15min before scanning with the ODyssey IRD scanner.
5 µl of 15μg/µl Solanum lycopersicum protein saturated in 8M urea were separated on 15% SDS-PAGE and blotted for 1hour to 0.2 µm nitrocellulose at 100V using wet transfer system. Blots were blocked with 0.5% cold fish gelatin for 1hr at room temp with agitation. Blot was incubated in the primary antibody (anti-H3) at a dilution of 1:2500 for an hour at RT with agitation. The blots were washed with 3X 15min TBS-TT at RT with agitation. Blots as incubated in the secondary antibody, fluorescent antibody (AS12 2460, Agrisera) 1:5000 dilution for 30min at RT with agitation and washed 1X with TBSTT for 15min, 1X with TBST for 15min before scanning with the ODyssey IRD scanner.
Courtesy of Dr. Betty Chung and Dr Zhengming Wang, University of Cambridge, United Kingdom

Immunocytochemical assays were performed according to the method described earlier (Rybaczek and Maszewski 2006). Excised apical parts of Vicia faba roots (1.5 mm long) were fixed for 45 min (18°C) in PBS-buffered 3.7% paraformaldehyde, washed several times with PBS and placed in a citric acid-buffered digestion solution (pH 5.0; 37°C for 45 min) containing 2.5% pectinase (Fluka), 2.5% cellulase (Onozuka R-10; Serva) and 2.5% pectoliase (ICN). After removing the digestion solution, root tips were washed 3 times in PBS, rinsed with distilled water and squashed onto Super Frost Plus glass slides (Menzel-Gläser). Air-dried slides were pretreated with PBS-buffered 5% BSA at 20°C for 50 min and incubated overnight in a humidified atmosphere (4°C) with rabbit antibody raised against H3 histone (Agrisera), dissolved in PBS containing 1% BSA (at a dilution of 1:50). Following incubation, slides were washed 3 times with PBS and incubated for 1 h (18°C) with Agrisera secondary goat anti-rabbit IgG DyLight®488 antibody (AS09 633, 1:1000). Nuclear DNA was stained with 4’,6-diamidino-2-phenyl-indole (DAPI, 0.4 μg/ml; Sigma-Aldrich). Following washing with PBS, slides were air dried and embedded in Vectashield Mounting Media for Fluorescence (Vector Laboratories). Observations were made using Optiphot-2 fluorescence microscope (Nikon) equipped with B-2A filter (blue light; λ ≈ 495 nm) for DyLight-conjugated antibodies and UV-2A filter (UV light; λ ≈ 365 nm) for DAPI. All images were recorded at exactly the same time of integration using DXM 1200 CCD camera.
Courtesy Dr. Dorota Rybaczek, Łódź University, Poland
Application examples: 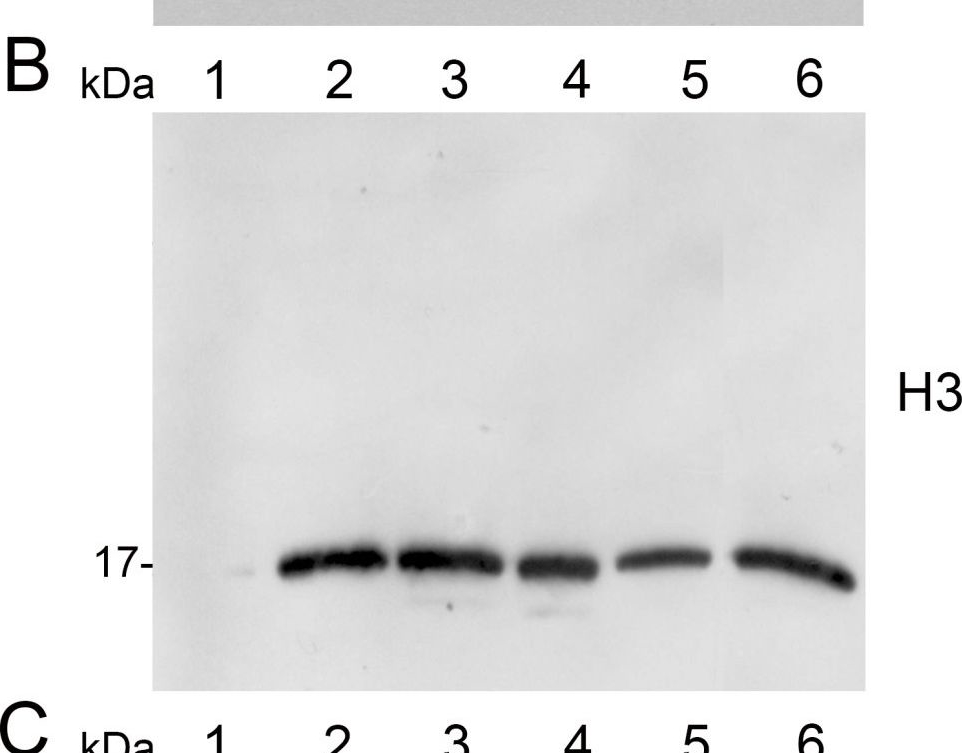
Reactant: Malus domestica (Apple)
Application: Western Blotting
Pudmed ID: 23886449
Journal: Plant Methods
Figure Number: 2B
Published Date: 2013-07-28
First Author: Sikorskaite, S., Rajamäki, M. L., et al.
Impact Factor: 5.139
Open PublicationAnalysis of protein fractions from different steps of nuclei isolation. The protein fractions obtained following the main steps of the procedure used for isolation of nuclei from leaves of apple are shown. Equal amounts of proteins (4 ?g) from each fraction were loaded on the gel. A) Lumenal-binding protein 2 (BiP2), B) histone H3 and C) plastocyanin (PC) were detected with specific antibodies by western blot analysis. D) Coomassie blue staining of the fractionated proteins separated by polyacrylamide gel electrophoresis. Lane 1: crude extract of proteins from homogenized leaf tissue; lane 2: resuspended pellet of the whole cell lysate obtained following treatment with Triton X-100 and centrifugation; lane 3: nuclei collected from the 60% Percoll layer (see nuclear fraction in Figure 1B); lane 4: the interface fraction of 60% Percoll and 2.5 M sucrose layers in the density gradient containing chloroplasts and unbroken cells (see 60% P/2.5 M S interface in Figure 1B); lane 5: sucrose layer of the density gradient (see 2.5 M sucrose layer in Figure 1B); lane 6: nuclei purified by centrifugation on a 35% Percoll cushion.
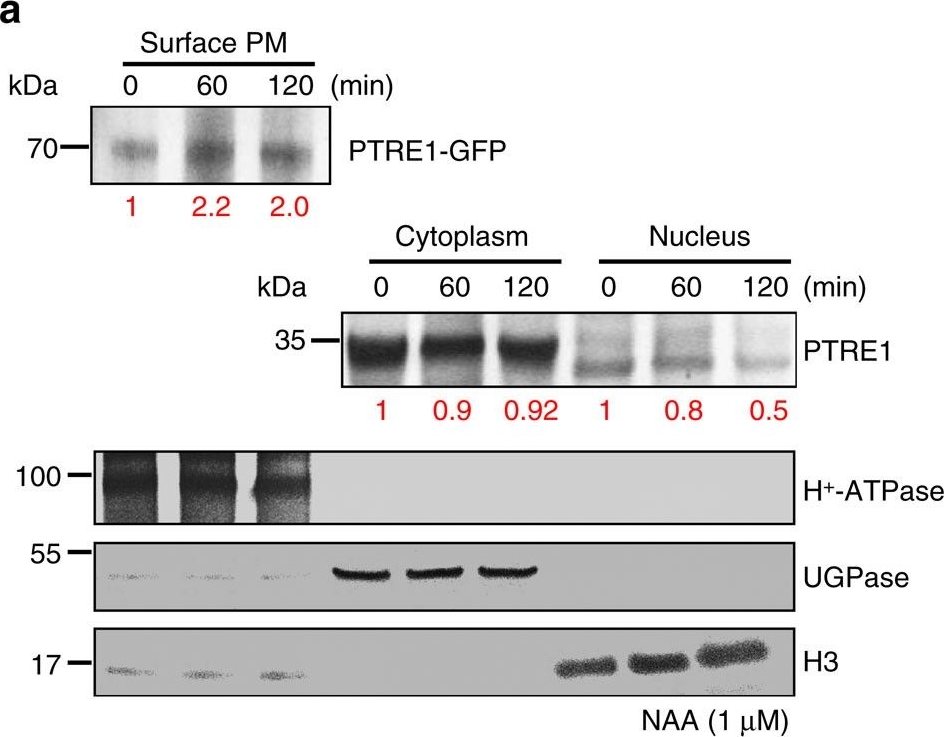
Reactant: Arabidopsis thaliana (Thale cress)
Application: Western Blotting
Pudmed ID: 27109828
Journal: Nat Commun
Figure Number: 6A
Published Date: 2016-04-25
First Author: Yang, B. J., Han, X. X., et al.
Impact Factor: 13.783
Open PublicationAuxin stimulates PTRE1 accumulation at the plasma membrane.(a) Western blot analysis showed that auxin treatment results in relatively more PTRE1 at the plasma membrane, but decreased levels in nucleus and cytoplasm. Arabidopsis seedlings expressing PTRE1-GFP were treated with NAA (1??M, 60 or 120?min) and then surface-exposed membrane proteins were analysed using PTRE1 antibody or plasma membrane marker H+-ATPase. Nucleus and cytoplasm fractions were prepared from Col seedlings and analysed by western blot using antibody against PTRE1, nuclear marker H3 or cytoplasm marker UGPase. The relative quantities of the proteins were calculated by using image pro plus and indicated. (b) A proposed model for how PTRE1 and TIR1 coordinate auxin responses regulating proteasome activity and Aux/IAA protein degradation. Under normal condition (with basal auxin levels), 26S proteasome activity is maintained by appropriate distribution of PTRE1 at the plasma membrane and in intracellular compartments. In response to auxin, auxin rapidly stimulates the association of TIR1 and Aux/IAA proteins resulting in degradation of Aux/IAAs (1). Later, auxin suppresses PTRE1 to inhibit proteasome activity (possibly through stimulating the accumulation of PTRE1 at plasma membrane, resulting in decreased intracellular and nuclear localization) and hence suppresses Aux/IAA protein degradation (2) to coordinate regulation of auxin responses, reflecting a mechanism for fine control of Aux/IAA homeostasis and auxin signalling.
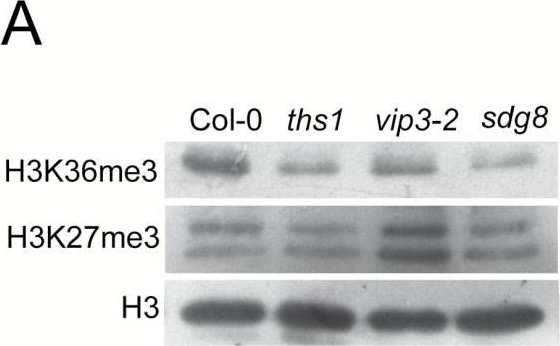
Reactant: Arabidopsis thaliana (Thale cress)
Application: Western Blotting
Pudmed ID: 28204553
Journal: J Exp Bot
Figure Number: 4A
Published Date: 2017-01-01
First Author: Jensen, G. S., Fal, K., et al.
Impact Factor: 6.088
Open Publicationths1 and vip3-2 mutants have altered global and genic histone H3 methylation patterns and fail to induce TCH3 and TCH4 in response to touch. (A) Chromatin extracts from cauline leaves of the indicated genotypes were separated by SDS-PAGE and detected with antibodies specific to H3K36me3 or H3K27me3. (B) Relative enrichment for H3K36me3 at the TCH3 and TCH4 loci in cauline leaf chromatin. Chromatin was isolated as in (A), then immunoprecipitated with the H3K36me3 antibody or without antibody and precipitated DNA amplified by qRT-PCR. Data presented is a percentage of the input, normalized to the S-ADENOSYLMETHIONINE SYNTHASE (SAM) gene and represented as a fold difference with the wild type non-treated sample. Error bars indicate the SEM for three biological replicates with two technical replicates each. (C) Quantitative reverse-transcriptase PCR analysis of TCH3 and TCH4 expression in response to touch in wild type and mutant seedlings. Seedlings were brushed for 2 minutes with a paintbrush according to the scheme shown in the top panel. RNA was prepared from the aerial tissues of treated and untreated plants and cDNA amplified with gene specific primers. The data was normalized to TUB4 (upper panels) or TCTP (lower panels) as reference genes and is presented as fold change compared to wild type non-treated samples. The error bars indicate SEM from three replicates. Asterisks mark significant difference from the untouched control P<0 05.
Reactant: Arabidopsis thaliana (Thale cress)
Application: Western Blotting
Pudmed ID: 28935922
Journal: Nat Commun
Figure Number: 2D
Published Date: 2017-09-21
First Author: Chen, P., Takatsuka, H., et al.
Impact Factor: 13.783
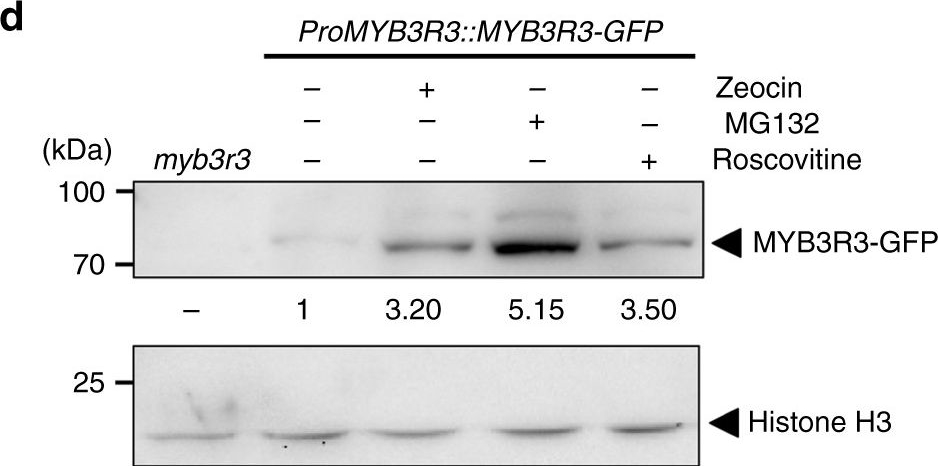
Open PublicationMYB3R3 accumulation in the root tip. a, c Confocal microscopy images of root tips. Five-day-old myb3r3 seedlings harbouring ProMYB3R3::MYB3R3-GFP were treated with 2??M zeocin for the indicated times a, or with 2??M zeocin, 50??M MG132 or 25??M roscovitine for 24?h c. Scale bars, 100??m. b, d Protein level of MYB3R3-GFP in roots. Ten-day-old myb3r3 seedlings harbouring ProMYB3R3::MYB3R3-GFP were treated with 5 or 10??M zeocin for 24 or 48?h b, or with 2??M zeocin, 50??M MG132 or 25??M roscovitine for 24?h d. Forty (b) or 20?µg (d) of total protein extracted from roots were subjected to immunoblotting using antibodies against GFP, ?-tubulin or histone H3. Protein extract from the myb3r3 mutant was used as a control (d). Relative levels of MYB3R3-GFP are expressed as the fold change, normalized with respect to the band of ?-tubulin or histone H3
Reactant: Arabidopsis thaliana (Thale cress)
Application: Western Blotting
Pudmed ID: 28935922
Journal: Nat Commun
Figure Number: 6B
Published Date: 2017-09-21
First Author: Chen, P., Takatsuka, H., et al.
Impact Factor: 13.783
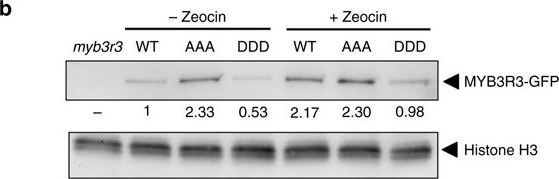
Open PublicationPhosphorylation state of MYB3R3 is associated with protein accumulation and zeocin response. a Confocal microscopy images of myb3r3 root tips harbouring ProMYB3R3::MYB3R3AAA-GFP or ProMYB3R3::MYB3R3DDD-GFP. Five-day-old seedlings were treated with or without 2??M zeocin, or with 50??M MG132, for 24?h. Scale bars, 100??m. b Protein level of MYB3R3-GFP in roots. Ten-day-old myb3r3 seedlings harbouring ProMYB3R3::MYB3R3-GFP (WT), ProMYB3R3::MYB3R3AAA-GFP (AAA) or ProMYB3R3::MYB3R3DDD-GFP (DDD) were treated with or without 2??M zeocin for 24?h, and 40?µg of total protein extracted from roots was immunoblotted with anti-GFP or anti-histone H3 antibodies. Protein extract from myb3r3 was used as a control. Relative levels of MYB3R3-GFP are expressed as the fold change, normalized with respect to the band of histone H3. c Eighteen-day-old seedlings of WT, myb3r3 and myb3r3 carrying ProMYB3R3::MYB3R3AAA-GFP or ProMYB3R3::MYB3R3DDD-GFP. Scale bars, 2?cm. d, e Root growth of WT, myb3r3 and myb3r3 carrying ProMYB3R3::MYB3R3-GFP, ProMYB3R3::MYB3R3AAA-GFP or ProMYB3R3::MYB3R3DDD-GFP. Five-day-old seedlings were transferred to medium with or without 2?µM zeocin, and root length was measured for 6 days d. Root length after 6 days of zeocin treatment is shown in e. Data are presented as mean?±?SD (n?>?30). Significant differences from WT were determined by Student’s t-test: *P?
Reactant: Arabidopsis thaliana (Thale cress)
Application: Western Blotting
Pudmed ID: 29180698
Journal: Nat Commun
Figure Number: 4B
Published Date: 2017-11-27
First Author: Huot, B., Castroverde, C. D. M., et al.
Impact Factor: 13.783
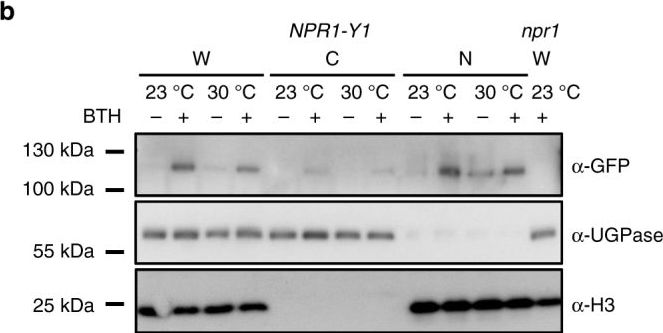
Open PublicationInduction of NPR1 nuclear localization is retained at 30?°C. a Representative confocal microscopy images of NPR1-YFP plants (n?=?4) 24?h after spraying with mock or BTH. Images are of YFP (yellow) alone or YFP overlaid on Brightfield (BF, grey-scale). Scale bar length represents 10?µm. b Western blots of whole-cell lysate (W), non-nuclear (C, cytosolic) and nuclear (N) enriched fractions isolated from NPR1-Y1 transgenic plants treated with mock (?) or BTH (+) solutions at 23?°C or 30?°C. Equal volumes (10?µl) of each protein sample were loaded and run in two separate 4–12 % gradient SDS-PAGE gels. Following transfer to PVDF membranes, one blot was probed with ?-GFP primary antibody to detect the NPR1-YFP protein while the other blot was cut in two and the upper portion probed with ?-UGPase (a cytosolic protein control) and the lower portion probed with ?-H3 (a nuclear protein control). Whole-cell lysate extracted from npr1 plants treated with BTH at 23?°C was used as the negative control for the NPR1-YFP band. All data are representative of three independent experiments

Reactant: Botrytis cinerea
Application: Western Blotting
Pudmed ID: 32098836
Journal: mSystems
Figure Number: 1E
Published Date: 2020-02-25
First Author: Li, H., Zhang, Z., et al.
Impact Factor: None
Open PublicationGeneration of ?bcactA mutants. (A) Replacement strategy for deletion of the bcactA gene. The 5? flank and 3? flank of bcactA were amplified and ligated into pLOB7 to flank the hygromycin resistance cassette to obtain the replacement vectors. Deletion mutants were generated by replacing the bcactA gene with the hygromycin resistance cassette through transformation of the protoplasts of the wild-type (WT) strain. (B) PCR diagnosis of ?bcactA mutants. PCR was performed using the primer pair bcactA-homo-up/HPH-det, to ensure that homologous recombination occurred at the target site. (C) Diagnosis of the homozygotes of ?bcactA mutants. PCR was performed with the primer pair bcactA-homo-up/bcactA-R-down, to ensure that the mutants were homozygous with no wild-type bands. (D) Southern blot analysis of WT and ?bcactA strains. The genomic DNA was digested with SacI and BamHI, separated in an agarose gel, and hybridized with a probe (a fragment on the hygromycin resistance cassette labeled with digoxigenin). Numbers represent different strains. (E) Immunoblotting analysis of BcactA protein in WT and ?bcactA strains. Total cytoplasmic proteins were extracted from 2-day-old mycelia cultured in PDB and immunoblotted with commercially available antibody to actin (Abmart; M20011). Histone 3 (H3) was used as a loading control.
Reactant: Arabidopsis thaliana (Thale cress)
Application: Western Blotting
Pudmed ID: 32210987
Journal: Front Plant Sci
Figure Number: 3A
Published Date: 2020-03-27
First Author: Fu, C., Liu, X., et al.
Impact Factor: 5.435
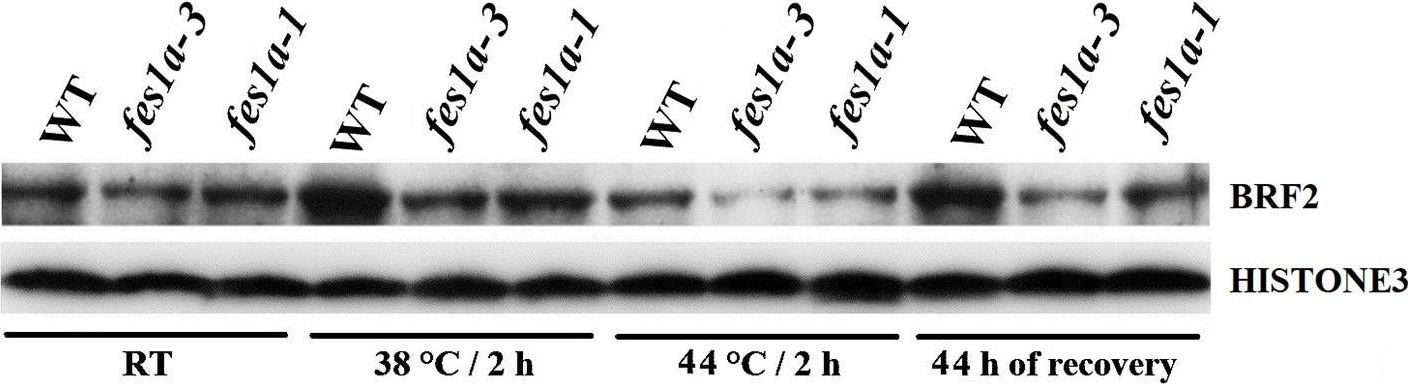
Open PublicationWestern blot results exhibiting the expression of BRF2 protein in 10-day-old seedlings of WT and fes1a mutants.
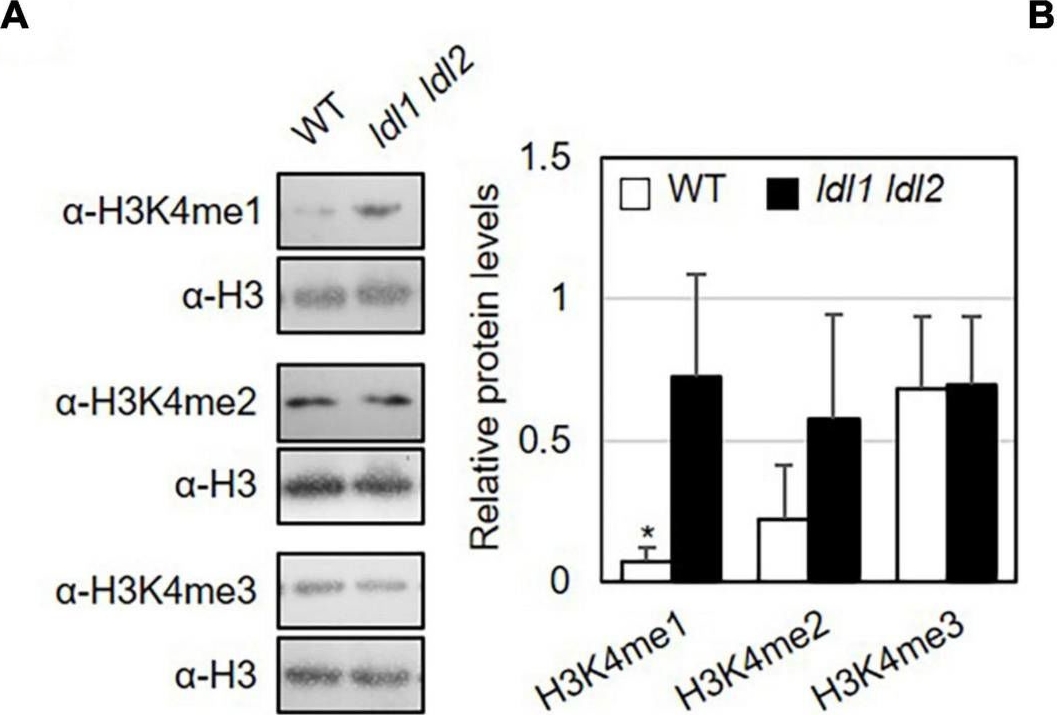
Reactant: Arabidopsis thaliana (Thale cress)
Application: Western Blotting
Pudmed ID: 34194459
Journal: Front Plant Sci
Figure Number: 5A
Published Date: 2021-07-02
First Author: Noh, S. W., Seo, R. R., et al.
Impact Factor: 5.435
Open PublicationMonomethylation of histone H3K4 is associated with the transcription of genes encoding transcription factors in Arabidopsis. (A) Abundance of H3K4me1, H3K4me2, and H3K4me3 marked proteins in WT and ldl1 ldl2 plants. The right panel shows modified histone H3 protein levels in Arabidopsis (average ± SD, *p < 0.05, Student’s t-test, n = 3). The results of all independent experiments are shown in Supplementary Figure 6. (B) Genomic DNA regions of gene-specific primers used in panels (C-E). (C-E) ChIP-qPCR to examine the enrichment of modified histone H3 proteins in the identified LDL1-target genes (C), transcription factors overrepresented in the ldl1 ldl2 mutants (D), and PR1 and FRK1 genes (E). Enrichment of Lys 4- mono-, Lys 4- di-, and Lys 4-trimethylated histone H3 protein at the chromatin regions of genes was measured in wild type (white bars) and ldl1 ldl2 (gray bars). The amount of DNA after ChIP was quantified by qPCR, and the means represent the average immunoprecipitation efficiencies (%) against total input DNA used. Each plot shows a representative of two independent biological replications (with similar results), and the data points are the average values of technical triplicates (average ± SEM, *p < 0.05, Student’s t-test, n = 3). The blue bars are the amplicon regions used for ChiP-qPCR, and the primer sequences used in this study are shown in Supplementary Table 2.
- Additional Information
-
Additional information: Cellular [compartment marker] of nucleoplasm, loading control antibody for Chlamydomonas reinhardtii
Additional information (application): Specific fluorescence in ICC has been observed for interphase nuclei as well as around centromer region (where Ser10 of histone H3 is phosphorylated) in mitotic chromosomes - Background
-
Background: Histone 3 (H3) located in nuclei, incorporated into chromatin. Present in nucleosome together with H2A, H2B and H4.
- Product Citations
-
Selected references: Sánchez-Vicente et al. (2026). Redox-mediated regulation of ABSCISIC ACID-INSENSITIVE 5 affects seed germination and seedling development in Arabidopsis. J Exp Bot . 2026 Feb 6:erag006. doi: 10.1093/jxb/erag006.
Grądzka et al.(2025). CHIP-seq and transcriptomics reveal a new role of circadian-regulated StBBX24 protein in potato reproduction. BMC Plant Biol. 2025 Dec 2;25(1):1702.doi: 10.1186/s12870-025-07811-0.
Chen et al. (2025). Short heat shock factor A2 regulates heat resistance and growth balance in Arabidopsis. Elife. 2025 Nov 3:13:RP99937. doi: 10.7554/eLife.99937.
Mohapatra et al. (2025). An Improved Method for Isolating Intact Chloroplasts from Different Plant Species. Journal of Plant Growth Regulation, doi.org/10.1007/s00344-025-11911-4.
Fal et al. (2025). CRISPR/dCas9-targeted H3K27me3 demethylation at the CUC3 boundary gene triggers ectopic transcription and impacts plant development. iScience . 2025 Apr 21;28(5):112475. doi: 10.1016/j.isci.2025.112475.
Supriya et al. (2025). Melatonin Bridges Timekeeping and Survival: Coordinating Diurnal Rhythms, Autophagy, and Energy Balance in Upland Cotton. Plant Stress, 100969.
Boussardon et al. (2025). The atypical proteome of mitochondria from mature pollen grains. Curr Biol . 2025 Jan 21:S0960-9822(24)01705-6. doi: 10.1016/j.cub.2024.12.037.
Martín-Merchán et al. (2024). Arabidopsis AGO1 N-terminal extension acts as an essential hub for PRMT5 interaction and post-translational modifications. Nucleic Acids Res . 2024 May 20:gkae387.doi: 10.1093/nar/gkae387.
Mosesso et al. (2024). Arabidopsis CaLB1 undergoes phase separation with the ESCRT protein ALIX and modulates autophagosome maturation. Nat Commun. 2024 Jun 19;15(1):5188. doi: 10.1038/s41467-024-49485-6.
Nguyen et al.(2024). The processed C-terminus of AvrRps4 effector suppresses plant immunity via targeting multiple WRKYs.J Integr Plant Biol. 2024 Jun 13.doi: 10.1111/jipb.13710.
Kimura and Kanegae (2024). A phytochrome/phototropin chimeric photoreceptor promotes growth of fern gametophytes under limited light conditions. Exp Bot . 2024 Apr 15;75(8):2403-2416.
Zhao et al. (2024). Psb28 protein is indispensable for stable accumulation of PSII core complexes in Arabidopsis.Plant J. 2024 May 26. doi: 10.1111/tpj.16844.
Ciesielska et al. (2024). S2P2-the chloroplast-located intramembrane protease and its impact on the stoichiometry and functioning of the photosynthetic apparatus of A. thaliana. Front Plant Sci. 2024 Mar 15:15:1372318. doi: 10.3389/fpls.2024.1372318.
Kim et al. (2024).Photoautotrophic cultivation of a Chlamydomonas reinhardtii mutant with zeaxanthin as the sole xanthophyll. Biotechnol Biofuels Bioprod. 2024 Mar 14;17(1):41. doi: 10.1186/s13068-024-02483-8.
Woo et al.(2024). Attenuation of phytofungal pathogenicity of Ascomycota by autophagy modulators. Nat Commun. 2024 Feb 29;15(1):1621.doi: 10.1038/s41467-024-45839-2.
Liu et al. (2023). RBPome identification in egg-cell like callus of Arabidopsis. Biol Chem. 2023 Sep 29;404(11-12):1137-1149.doi: 10.1515/hsz-2023-0195.
Hyun et al. (2023). Functional demonstration of Aureochrome 1a proteasomal degradation after blue light incubation in the diatom Phaeodactylum tricornutum. J Plant Physiol. 2023 Dec 1:292:154148. doi: 10.1016/j.jplph.2023.154148
Chih-Cheng Lin and others, SUB1A-1 anchors a regulatory cascade for epigenetic and transcriptional controls of submergence tolerance in rice, PNAS Nexus, 2023;, pgad229.
Ji et al (2023) The thioesterase APT1 is a bidirectional-adjustment redox sensor. Nat Commun. 2023 May 17;14(1):2807. doi: 10.1038/s41467-023-38464-y.
Park et al. (2023) Manavella PA, Yun DJ. The HOS15-HDA9 complex associates with HYL1 to modulate miRNA expression in response to ABA signaling. Plant Cell. 2023 May 17:koad132. doi: 10.1093/plcell/koad132.
Garay et al (2023) Perinatal Protein Restriction Impacts Nuclear O-GalNAc Glycosylation in Cells of Liver and Brain Structures of the Rat. J Nutr. 2023 Apr;153(4):979-987. doi: 10.1016/j.tjnut.2023.02.036.
Singh D, Datta S. BBX30/miP1b and BBX31/miP1a form a positive feedback loop with ABI5 to regulate ABA-mediated postgermination seedling growth arrest. New Phytol. 2023 Jun;238(5):1908-1923. doi: 10.1111/nph.18866.
Boussardon C, Carrie C, Keech O. Comparing plastid proteomes points towards a higher plastidial redox turnover in vascular tissues than in mesophyll cells. J Exp Bot. 2023 Apr 7:erad133. doi: 10.1093/jxb/erad133.
Hu et al. (2022) Bacterial effectors manipulate plant abscisic acid signaling for creation of an aqueous apoplast. Cell Host Microbe. 2022 Apr 13;30(4):518-529.e6. doi: 10.1016/j.chom.2022.02.002. Epub 2022 Mar 4. PMID: 35247331.
Liang et al. (2022). Arabidopsis RBV is a conserved WD40 repeat protein that promotes microRNA biogenesis and ARGONAUTE1 loading. Nat Commun. 2022 Mar 8;13(1):1217. doi: 10.1038/s41467-022-28872-x. PMID: 35260568; PMCID: PMC8904849.
Gomez et al. (2022) Phosphatidylinositol-4-phosphate controls autophagosome formation in Arabidopsis thaliana. Nat Commun. 2022 Jul 28;13(1):4385. doi: 10.1038/s41467-022-32109-2. PMID: 35902598; PMCID: PMC9334301.
Skalický et al. (2023). Fluorescence-activated multi-organelle mapping of subcellular plant hormone distribution. Plant J. 2023 Dec;116(6):1825-1841.doi: 10.1111/tpj.16456. Epub 2023 Sep 8.
Farago et al. (2022) Small paraquat resistance proteins modulate paraquat and ABA responses and confer drought tolerance to overexpressing Arabidopsis plants. Plant Cell Environ. 2022 Jul;45(7):1985-2003. doi: 10.1111/pce.14338. Epub 2022 Apr 29. PMID: 35486392; PMCID: PMC9324991.
Margaritopoulou et al (2021) Enriched HeK4me3 marks at Pm-0 resistance-related genes prime courgette against Podosphaera xanthii. Plant Physiol. 2021 Sep 21:kiab453. doi: 10.1093/plphys/kiab453. Epub ahead of print. Erratum in: Plant Physiol. 2021 Nov 11;: PMID: 34597395.
Perlaza (2021). Organelle Size and Quality Control in Chlamydomonas Reinhardtii. UCSF. ProQuest ID: Perlaza_ucsf_0034D_12217. Merritt ID: ark:/13030/m5257z1d. Retrieved from https://escholarship.org/uc/item/1jg3874h
Sun et al. (2021) The epigenetic factor FVE orchestrates cytoplasmic SGS3-DRB4-DCL4 activities to promote transgene silencing in Arabidopsis. Sci Adv. 2021 Aug 4;7(32):eabf3898. doi: 10.1126/sciadv.abf3898. PMID: 34348894; PMCID: PMC8336953.
Chen et al. (2019). Phalaenopsis LEAFY COTYLEDON1-Induced Somatic Embryonic Structures Are Morphologically Distinct From Protocorm-Like Bodies. Front Plant Sci. 2019 Nov 29;10:1594. doi: 10.3389/fpls.2019.01594. PMID: 31850050; PMCID: PMC6896055.
Perlaza et al. (2019). The Mars1 kinase confers photoprotection through signaling in the chloroplast unfolded protein response. Elife. 2019 Oct 15;8. pii: e49577. doi: 10.7554/eLife.49577. (immunofluorescence)
Dalmadi et al. (2019). AGO-unbound cytosolic pool of mature miRNAs in plant cells reveals a novel regulatory step at AGO1 loading. Nucleic Acids Res. 2019 Aug 8. pii: gkz690. doi: 10.1093/nar/gkz690.
Barua et al. (2019). Dehydration-responsive nuclear proteome landscape of chickpea (Cicer arietinum L.) reveals phosphorylation-mediated regulation of stress response. Plant Cell Environ. 2019 Jan;42(1):230-244. doi: 10.1111/pce.13334.
Du et al (2019). Proteomic identification of lipid-bodies-associated proteins in maize seeds. Acta Physiologiae Plantarum, May 2019, 41:70
Lai et al. (2018). Salicylic acid-independent role of NPR1 is required for protection from proteotoxic stress in the plant endoplasmic reticulum. Proc Natl Acad Sci U S A. 2018 May 29;115(22):E5203-E5212. doi: 10.1073/pnas.1802254115.
Wang et al. (2018). Degradation of unmethylated miRNA/miRNA*s by a DEDDy-type 3' to 5' exoribonuclease Atrimmer 2 in Arabidopsis. Proc Natl Acad Sci U S A. 2018 Jul 10;115(28):E6659-E6667. doi: 10.1073/pnas.1721917115.
Hartmann et al. (2018). Subcellular Compartmentation of Alternatively Spliced Transcripts Defines SERINE/ARGININE-RICH PROTEIN30 Expression. Plant Physiol. 2018 Apr;176(4):2886-2903. doi: 10.1104/pp.17.01260.
Duan et al. (2017). A Lipid-Anchored NAC Transcription Factor Is Translocated into the Nucleus and Activates Glyoxalase I Expression during Drought Stress. Plant Cell. 2017 Jul;29(7):1748-1772. doi: 10.1105/tpc.17.00044. (Nicotiana benthamiana)
Rihan et al. (2017). An analysis of the development of cauliflower seed as a model to improve the molecular mechanism of abiotic stress tolerance in cauliflower artificial seeds. Plant Physiol Biochem. 2017 Jul;116:91-105. doi: 10.1016/j.plaphy.2017.05.011.
Shin et al. (2017). The metabolic sensor AKIN10 modulates the Arabidopsis circadian clock in a light-dependent manner. Plant Cell Environ. 2017 Jan 5. doi: 10.1111/pce.12903.
Correa-Galvis et al. (2016). Photosystem II Subunit PsbS Is Involved in the Induction of LHCSR Protein-dependent Energy Dissipation in Chlamydomonas reinhardtii. J Biol Chem. 2016 Aug 12;291(33):17478-87. doi: 10.1074/jbc.M116.737312.
Castellano et al. (2016). A pathogenic long noncoding RNA redesigns the epigenetic landscape of the infected cells by subverting host Histone Deacetylase 6 activity. New Phytol. 2016 Sep;211(4):1311-22. doi: 10.1111/nph.14001. Epub 2016 May 12.
Ghandi et al. (2016). Tomato yellow leaf curl virus infection mitigates the heat stress response of plants grown at high temperature. Sci Rep. 2016 Jan 21;6:19715. doi: 10.1038/srep19715.
Gorovits et al. (2016). Tomato yellow leaf curl virus confronts host degradation by sheltering in small/midsized protein aggregates. Virus Res. 2016 Feb 2;213:304-13. doi: 10.1016/j.virusres.2015.11.020. Epub 2015 Dec 1
Giaume et al. (2023) Two florigens and a florigen-like protein form a triple regulatory module at the shoot apical meristem to promote reproductive transitions in rice. Nat Plants. 2023 Apr;9(4):525-534. doi: 10.1038/s41477-023-01383-3.
Lan et al (2023) UPL5 modulates WHY2 protein distribution in a Kub-site dependent ubiquitination in response to [Ca2+]cyt-induced leaf senescence - Protocols
-
Agrisera Western Blot protocol and video tutorials
Protocols to work with plant and algal protein extracts
Agrisera Educational Posters CollectionPreparation of cytosolic and nuclear protein fractions
1. Prepare protoplasts from 50 ml Arabidopsis thaliana cell culture according to the protocol of PEG transfection.
2. Resuspend protoplasts in 10 ml GH buffer and keep the solution on ice for 10 min.
GH buffer: 100mM glycine
0.1% Hexylene glycol
0.37M (4.7% w/v) saccharose
0.3mM Spermine
1.0mM Spermidine
pH 8.3 with Ca(OH)2
3. To release nuclei add Triton X100 to a final concentration of 0.1%. Pipetting gently up and down several
times with a plastic pipette might be necessary to lyse cells.
4. After 5min sediment nuclei by centrifugation at 1000 xg for 15 min at 4°C. Save supernatant as the
cytoplasmic fraction. Wash the pelleted nuclei two times with GHT (GH+0.1% TX100) then finally
resuspended in a suitable volume of extraction buffer + protease inhibitors.Courtesy Dr. Laszlo Bako, Umeå Plant Science Centre
- Reviews:
-
Peter Ma | 2023-04-10This H3 antibody worked very well and showed no background with our samples. We used 1:5000 dilution of H3 for total Arabidopsis protein extractions and 1:10000 dilution for secondary antibody.KAILI XIE | 2021-05-12This H3 antibody worked well in western blot with a dilution of 1:5000 and the signal is very strong. If you want use it to do the ChIP assay, maybe you need to do the test firstly.Kaili Xie | 2021-05-06This H3 antibody worked really well in ChIP and western blot in our hands. The dilution for western blot is 1:5000 and signal is very strong!INMACULADA SÁNCHEZ-VICENTE | 2019-10-10We used H3 antibody with nuclear protein extracts from Arabidopsis at a dilution of 1:5000 and it gave very good result. I recomend this product.Sidona | 2011-11-16We have used H3 antibody with nuclear protein extracts from Nicotiana and Malus plants at a dilution of 1:10000 and it gave very strong signal. The same dilution could be re-used at least 3 times.Rena Gorovits | 2011-09-20strong signal in nuclear fraction of tomato protein extractsDr. Leonardo Magneschi | 2011-06-29I tested this antibody for WB on Chlamydomonas reinhardtii whole cells and nuclei-enriched protein samples at a dilution of 1:5000 (see figure in the ''info'' section of this antibody). The antibody has high affinity for the Chlamydomonas H3 protein and generates a good signal after short time exposures with standard ECL.
Accessories
AS09 602 | Clonality: Polyclonal | Host: Goat | Reactivity: Rabbit IgG (H&L)
AS09 607 | Clonality: Polyclonal Host: Goat Reactivity: Rabbit IgG (H&L)